II. Système GFP (Green Fluorescent Protein)
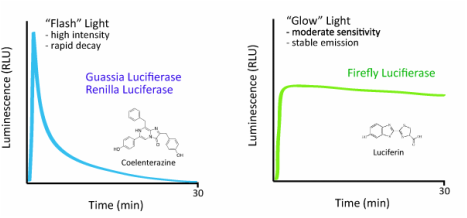
Dans ce système, l'équivalent de la luciférine (molécule utilisée pour émettre de la lumière) porte un nom propre : la coelenterazine. Contrairement à la luciférine, cette molécule permet une émission de lumière plus intense mais de courte durée, idéale pour la répulsion. La bioluminescence est intracellulaire. Ce mécanisme est utilisé par l'anémone Renilla reniformis et la méduse Aequorea victoria.
Lors de la réaction qui permet l'émission de lumière, la coelenterazine va s'associer avec de la luciférase qui va catalyser sa réaction avec du dioxygène. Il se formera du dioxyde de carbone et une molécule excitée: la coelenteramide qui émettra un rayonnement de longueur d’onde de 485nm (bleu) en se déséxcitant.
Coelenterazine + O2 + Luciférase ---> Coelenteramide* + CO2 + h𝝂 ( * = molécule excitée)
Dans ce système, l'équivalent de la luciférine (molécule utilisée pour émettre de la lumière) porte un nom propre : la coelenterazine. Contrairement à la luciférine, cette molécule permet une émission de lumière plus intense mais de courte durée, idéale pour la répulsion. La bioluminescence est intracellulaire. Ce mécanisme est utilisé par l'anémone Renilla reniformis et la méduse Aequorea victoria.
Lors de la réaction qui permet l'émission de lumière, la coelenterazine va s'associer avec de la luciférase qui va catalyser sa réaction avec du dioxygène. Il se formera du dioxyde de carbone et une molécule excitée: la coelenteramide qui émettra un rayonnement de longueur d’onde de 485nm (bleu) en se déséxcitant.
Coelenterazine + O2 + Luciférase ---> Coelenteramide* + CO2 + h𝝂 ( * = molécule excitée)
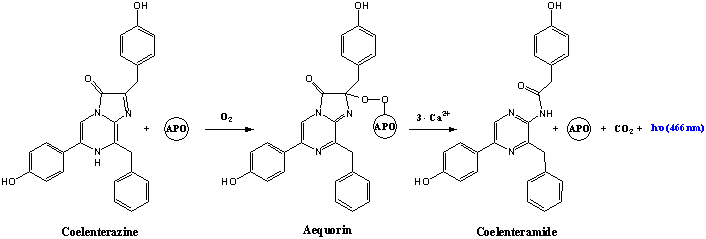
Dans la plupart des systèmes de bioluminescence, la rencontre de luciférine, luciférase et dioxygène produit imédiatement de la lumière. Cependant, la méduse a. victoria possède un système différent. Elle a une enzyme nomée apoaeqorine (APO sur le shéma ci-dessous) qui remplace la luciférase et est capable d'associer de la coelenterazine (équivalent de la luciférine) avec du dioxygène sans émission de lumière. Cette réaction intermédiaire est stable et forme une molécule nomée aeqorine qui, elle, émet un rayonnement d'environ 460nm en rencontrant des ions Ca2+. L' avantage pour cette méduse est que cette dernière réaction peut se faire encore plus rapidement.
coelenterazine + apo-aeqorine + O2 ---> aequorine + Ca2+ ---> apo-aequorine + CO2 + coelenteramide* + h𝝂
 Organisation de la protéine GFP de L'aequorea victoria
Organisation de la protéine GFP de L'aequorea victoria
Dans les deux cas, une protéine capable d’émettre de la lumière, la GFP va jouer un rôle important dans l' émission de lumière. Cette protéine possède un chromophore (groupe d'atomes qui lui permet d’émettre un rayonnement coloré) très similaire à une partie du chromophore de la coelenterazine. La GFP reste cependant stable car ce chromophore se trouve au centre de la protéine et est protégé par le reste des acides aminés. La coelenteramide sous état excité va se rapprocher de ce chromophore et permettre un transfert d’énergie.
Transfert d’énergie
Lors d'un transfert d’énergie entre molécules fluorescentes, une molécule excitée (le donneur) va transférer son énergie à une autre molécule également capable d’émettre de la lumière (l’accepteur). Pour que cela se produise il faut que les molécules se trouvent très proches (moins de 50nm) l’une de l’autre et que le spectre d’émission du donneur recouvre entièrement ou partiellement le spectre d’absorption de l’accepteur.
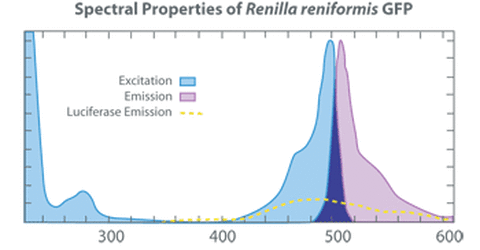
Dans ce cas, la coelenteramide est le donneur et la GFP l’accepteur, on remarque bien que le spectre d`absorbance de la GFP (en rose) recouvre une partie du spectre d`émission de la coelenteramide (en bleu clair) chez la renilla reniformis (dont le spectre d'émission est très proche de celui de la aequorea victoria). Le transfert d’énergie est donc possible et la GFP va émettre un rayonnement de 510nm (vert). L'avantage pour ces espèces est que la lumière verte est plus répulsive que la lumière bleue.
Le système GFP n’est pas la seule variante du mécanisme utilisé pour émettre de la lumière biologique, mais comme l’utilisation d’une GFP est très pratique puisqu’elle permet de modifier le spectre de la lumière émise par la réaction chimique afin qu’elle soit plus perceptible dans le milieu où se trouve l’émetteur, ce système est souvent combiné à d’autres mécanisme d’émission de lumière.
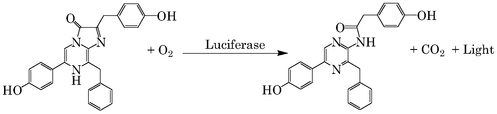
Il existe également d'autres mécanismes de bioluminescence qui associent des séries de réactions complexes comme dans le cas du système ATP.
